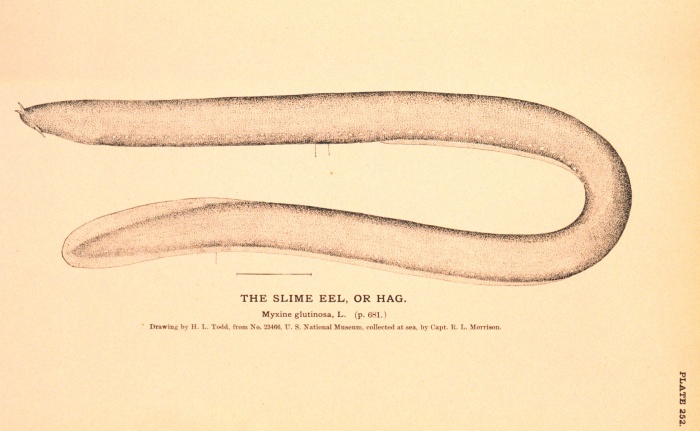
The Hagfish or Slime Eel, a modern Craniate related to the
Conodont-bearing
organism known from Cambrian-Triassic age rocks.
Myxine glutinosa L., figb0523, Historic NMFS Collection, NOAA
Invertebrate Paleontology Lab #12
Hemichordates
(Graptolites) and Craniates (Conodonts)
Click on the lab title to see the University of
California
Museum of Paleontology web page
Read BEFORE Coming to Lab: Benton & Harper, p. 409-423, 430-435, and 208-218
Introduction
This week
we
will explore two fascinating fossil groups that provide well known
index
fossils in biostratigraphy, and yet are still somewhat mysterious
organisms.
At this point in our march through the invertebrate phyla, we have
reached
the groups known as the Hemichordate Phylum and at long last, the
Chordate
Phylum, which includes the vertebrates. Both the graptolites,
which are hemichordates, and the conodonts, which are remains
of
members of the craniate subphylum in the Chordates, were in use as
major
tools in biostratigraphy long before systematists had clearly
identified
what they were!
The HEMICHORDATE PHYLUM (represented in the
fossil record by graptolites)
The Hemichordate
Phylum ("half-chordates") are a group of invertebrates that have
certain
features also shared by the Chordate Phylum, including a pharynx
("throat")
with multiple openings, a dorsal nerve chord and a
ventral
blood vessel, but unlike the Chordates, they have no
notochord.
A notochord is a stiff supporting rod that runs dorsally, just
above
the dorsal nerve chord in the Chordates, and is part of the vertebral
column
in the vertebrates. Instead of a notochord, Hemichordates have a
tube that is developed from the gut (intestines), that may be a
precursor
of the notochord. Living hemichordates include the acorn worms and the
pterobranchs. The fossil hemichordates of interest to us this
week
are the Graptolites (Late Cambrian-Pennsylvanian, with most
occurrences
in the Ordovician-Silurian).
Why the fuss about
graptolites?
They are ideal index fossils:
abundant, rapidly
evolving, easily fossilized (carbonized), wide ranging (planktonic life
style), distinctly identifiable.
They are the basis for the biostratigraphy of the Ordovician
and Silurian rocks.
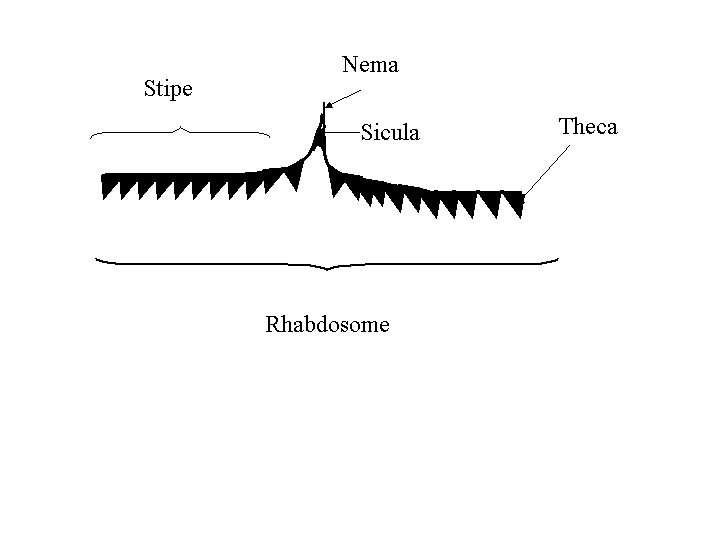
What are they?
Graptolites were colonial filter feeding organisms
that
apparently floated in surface ocean water in a range of depths. A
colony (called a rhabdosome) consisted of bunches of
branching
structures called stipes that are covered with tiny tubes or
cups
called thecae. In each cup was an individual zooid.
The base of the rhabdosome is the sicula, which is the first
zooid
theca that founded the colony. Sometimes there is a long stem
emerging
from the sicula called the nema, that probably attached the
stipe
to a floating structure. The branches or stipes are often found
fossilized
as carbonized remains. Graptolites did not produce a shell or calcium
phosphate
endoskeleton. They were simply tubes and cups of chitin,
connected
into a branching colony and attached to a float of some kind.
Drawing Graptolites
Examine the Graptolites in the teaching
collection, and
draw a specimen, labelling the stipe, sicula, nema and theca. |
The CHORDATE PHYLUM (represented by the Vertebrates,
but also by the CRANIATES, which left tooth-like structures called CONODONTS)
The Chordate
Phylum includes several subphyla (such as the vertebrates),
and
one of those subphyla is that of the Craniates-those chordate
animals
that have a brain and clearly defined head region, although no
vertebra:
instead, they have an internal skeleton of cartelage to support
them.
Living craniates today include the lamprey eel and the hagfish
or "slime eel" (see picture at top of page). Our interest in such
craniates
as the hagfish comes from the kind of teeth we find in them.
Hagfish
have tiny microscopic tooth-like structures on their tongues which they
use in a kind of rasping process. The hagfish tooth structures
are
very similar to microfossil structures found in Cambrian-Triassic
rocks called Conodonts. However, unlike true teeth,
these tiny structures were apparently always covered in tissue:
that
is, they show no wear. They are composed of calcium phosphate, a
typical bone material, and have cellular bone material in them (Sansom
et al., 1992). These two features imply that they are remains
from
some chordate animal. Unfortunately, the animals that produced
these
structures have never been definitely identified-the closest link has
been
to a hagfish-like fossil from the Lower Silurian in Wisconsin
(Mjickulic
et al., 1985; Smith et al., 1987).
Since we aren't all that
sure
what animal they came from, how are conodonts used?
Conodonts are extremely useful
as
biostratigraphic tools in correlation, because they are great index
fossils.
They are very abundant in Paleozoic rocks, and widely distributed,
rapidly
evolving, and easily identified.
Conodonts change color with
thermal
metamorphism. As the sedimentary rocks that enclose them are
subjected
to increasing temperature, conodonts will change from pale yellow to
darker
brown to black. This means they can be used to judge the
temperatures
at which a potential hydrocarbon reserve has been subjected to, and
ultimately,
if those rocks are potential targets for hydrocarbon recovery.
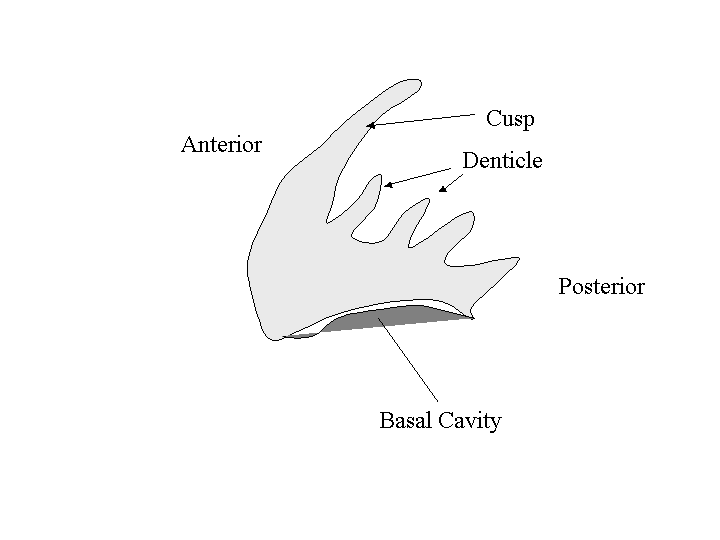
Drawing Conodonts
| Examine the conodonts in the teaching
collection and
draw the contents on any two slides, labelling the anterior, posterior,
cusp,denticle, and basal cavity. |

http://www.afsc.noaa.gov/race/media/photo_gallery/photos/Myxinidae/hagbill.jpg
Living hagfish-this species is Eptatretus deani, being dealt with
bravely by an employee of the Alaska Fisheries Science Center-NOAA
References
Mjickulic, D.G., Briggs, D.E., and Kluessendorf, J.,
1985.
A Silurian soft-bodied biota, Science, 228-714-717.
Sansom, I.J. and others, 1992. Presence of the
earliest
vertebrate hard tissues in conodonts. Science, 256, p
1308-1311.
Smith, M.P., Briggs, D.E., and Aldridge, R.J.,
1987.
A conodont animal from the lower Silurian of Wisconsin, U.S.A. and the
apparatus architecture of panderodontid conodonts, p 91-104 in
Aldridge,
R.J., (ed.) Palaeobiology of Conodonts, Ellis Horwood, Chichester.